ヒアルロン酸関節内注射の作用機序から適応疾患、分子量による効果の違いまで医学的エビデンスに基づいて解説。変形性関節症治療における最適な投与タイミングや期間について、最新の研究結果を踏まえた臨床応用とは?
骨芽細胞の働きと骨代謝における調節機構
骨芽細胞は骨形成の中心的役割を担う細胞です。本記事では、骨芽細胞の分化過程、骨基質合成メカニズム、破骨細胞との相互作用について解説します。最新の骨再生医療への応用も解説しますが、骨芽細胞研究の今後の展望はどうなるのでしょうか?

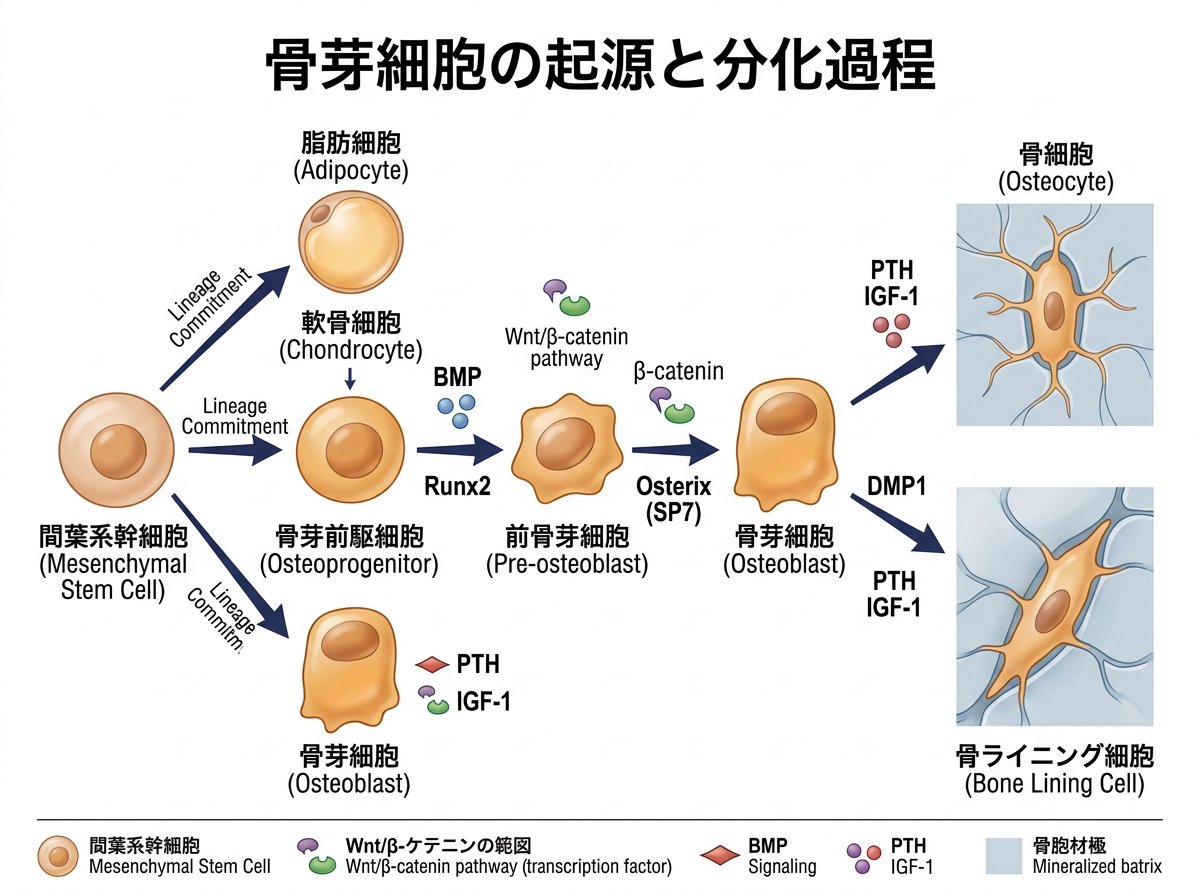
骨芽細胞は間葉系幹細胞から分化する細胞で、骨形成において中心的な役割を担っています。間葉系幹細胞は骨芽細胞だけでなく、軟骨細胞、脂肪細胞、筋肉細胞などの様々な細胞へと分化する多能性を持っています。骨芽細胞への分化過程では、特定の転写因子や成長因子が重要な役割を果たしています。
生体内では、間葉系幹細胞は骨膜直下に線維芽細胞様の形態で存在し、徐々に骨表面へと移動しながら分化を進めていきます。最終的に骨表面に一列に並んだ立方体様の骨芽細胞へと形質を変えます。この分化過程は厳密に制御されており、骨形成に必要な様々な遺伝子の発現が段階的に調整されています。
骨の発生には主に2つの経路があります。
この2つの異なる経路で形成される骨芽細胞は、基本的な性質が類似していると考えられています。
骨芽細胞への分化には複数の転写因子が関与していますが、特に転写因子Sp7(別名Osterix)は骨形成に必須であることが知られています。Sp7を欠損したマウスでは骨形成が著しく障害されることが報告されており、骨芽細胞の分化と機能において重要な役割を果たしています。
骨芽細胞の主要な機能は、骨基質の合成と石灰化の誘導です。骨芽細胞はまず、骨の有機成分の主体であるタイプIコラーゲンを大量に産生・分泌します。このコラーゲン線維が骨の基本的な枠組みを形成し、後の石灰化プロセスの足場となります。
コラーゲン以外にも、骨芽細胞は以下のような様々な非コラーゲン性タンパク質を産生します。
これらの分子は骨基質の構造形成や石灰化プロセスの調節に重要な役割を果たしています。
石灰化の過程では、骨芽細胞は「基質小胞(matrix vesicle)」と呼ばれる細胞外小胞構造物を骨基質に分泌します。この基質小胞内でリン酸カルシウムの初期結晶が形成され、その後結晶成長が進行してヒドロキシアパタイト[Ca10(PO4)6(OH)2]として骨基質に沈着します。
興味深いことに、カルシウムは体内で最も多い無機質であり、体重の約2%を占めています。そのうち99%は骨に存在しているのです。
活発に骨基質合成を行う骨芽細胞(活性型骨芽細胞または成熟型骨芽細胞と呼ばれる)は、発達した粗面小胞体やゴルジ体を持ち、これらの細胞内小器官は活発なタンパク質合成を反映しています。特にトランスゴルジネットワークの空胞内には、合成途中のコラーゲン線維などを観察することができます。
骨芽細胞は骨基質の形成を進めるにつれて、自らが産生した骨基質の中に埋め込まれていき、やがて骨細胞へと分化します。この骨細胞は骨芽細胞が静止した状態となり、骨質の中に閉じ込められた状態です。このように、骨芽細胞は骨基質の合成と石灰化を導くだけでなく、自らが骨細胞へと変化することで、骨組織の一部となっていくのです。
骨の細胞における組織学的・微細構造学的知見(日本顕微鏡学会)
骨組織は常に動的な状態にあり、骨芽細胞による骨形成と破骨細胞による骨吸収のバランスによって骨量が維持されています。この絶妙なバランスが「骨リモデリング(骨代謝回転)」と呼ばれるプロセスを支えています。
破骨細胞は骨髄または血液中に存在するマクロファージ系の造血細胞から形成される多核巨細胞で、石灰化した骨組織を吸収する唯一の細胞です。興味深いことに、マクロファージは体内の多くの場所に存在するにもかかわらず、破骨細胞は骨組織にのみ存在します。この現象の鍵を握っているのが骨芽細胞なのです。
骨芽細胞は破骨細胞の形成と機能を制御する重要な役割を担っています。具体的には、以下の因子を産生することで破骨細胞の分化を調節しています。
このRANKL/OPGのバランスが骨代謝の調節において非常に重要です。例えば、骨粗鬆症ではこのバランスが乱れ、RANKL優位となることで破骨細胞の活性が高まり、骨吸収が促進されます。
最近の研究では、骨芽細胞は小胞を放出・取り込みすることで互いに連携し合っていることが明らかになっています。大阪大学の研究グループは、骨芽細胞が骨表面で集団を形成して存在し、小胞を介してコミュニケーションを取りながら骨代謝を制御していることを報告しています。
また、興味深い発見として、骨芽細胞が産生するSema3Aというたんぱく質があります。これは骨吸収の抑制と骨形成の促進の双方を制御する二重の働きを持ち、適切な骨量維持に重要な役割を果たしています。
| 因子 | 産生細胞 | 主な作用 |
|---|---|---|
| M-CSF | 骨芽細胞 | 破骨細胞前駆細胞の生存・増殖促進 |
| RANKL | 骨芽細胞 | 破骨細胞分化の促進 |
| OPG | 骨芽細胞 | RANKL阻害による破骨細胞形成抑制 |
| Sema3A | 骨芽細胞 | 破骨細胞抑制と骨形成促進の二重作用 |
このように、骨芽細胞は単に骨を作るだけでなく、破骨細胞との複雑な相互作用を通じて骨のホメオスタシス(恒常性)を維持する中心的な役割を果たしています。
骨芽細胞の分化と機能において、転写因子Sp7(Osterix)は極めて重要な役割を果たしています。Sp7は骨形成に必須の転写因子であり、Sp7を欠損したマウスでは骨形成が著しく障害され、内軟骨性骨化と膜性骨化の両方が阻害されることが知られています。
Sp7はSpファミリーに属する転写因子で、従来は高度に保存されたジンクフィンガードメインを介してGCボックスと呼ばれるDNA配列に結合すると考えられていました。しかし、ChIP-seq法による解析から意外な発見がありました。骨芽細胞におけるSp7の結合モチーフにはGCボックスは有意に存在せず、むしろホメオボックス転写因子との結合配列を含むATリッチモチーフが大部分を占めていたのです。
この発見により、Sp7の作動様式に関する新たなモデルが提唱されました。Sp7は骨芽細胞において、ジンクフィンガードメインを介してホメオボックス転写因子(Dlx3、Dlx5、Dlx6など)と転写複合体を形成し、ATリッチモチーフに作用することにより遺伝子発現を制御すると考えられています。
この作動様式は、Spファミリーの他のメンバーとは全く異なるものであり、ChIP-seq法による解析によってはじめて明らかにされた生命現象の一例です。従来の試験管内(in vitro)での解析では検出することが困難だったこの現象は、骨形成メカニズムに新たな視点をもたらしています。
Sp7の標的となるゲノム領域には。
などが存在し、Sp7により制御される転写ネットワークは骨芽細胞の分化と機能に不可欠です。
また、生物種間の比較ゲノム解析により、Sp7の出現は骨組織をもつ脊椎動物の進化と密接に関連している可能性が示唆されています。このことは、Sp7が骨格系の進化において重要な役割を果たしてきたことを示唆しており、骨芽細胞の機能と骨形成のメカニズムを理解する上で重要な知見です。
骨再生医療は、外傷や疾患などで損傷した骨組織を修復・再生するための革新的なアプローチとして注目されています。この分野において、骨芽細胞は中心的な役割を果たしています。
骨再生医療の基本的な考え方は、人工的に作られた細胞や組織を用いて、損傷した組織や器官を修復・再生することです。骨の場合、その形成を担う骨芽細胞の移植が理想的なアプローチとなります。しかし、臨床応用においては、十分な量と質の骨芽細胞を確保することが大きな課題となっています。
骨芽細胞を直接移植する方法では、患者自身の骨髄から骨芽細胞を採取・培養する必要がありますが、これには手術的な負担やリスクが伴います。そのため、近年は以下のような代替アプローチが進展しています。
特に注目されているのが、骨芽細胞が産生する細胞外小胞の利用です。これらの小胞には様々な成長因子やmicroRNAが含まれており、骨形成を促進する効果があります。細胞そのものではなく小胞を利用することで、免疫拒絶反応のリスクを低減できる可能性があります。
また、近年開発が進んでいるのが、特定の分子シグナルを標的とした薬剤です。例えば、骨芽細胞の活性化や破骨細胞の抑制を促すSema3Aのような分子を活用した創薬研究も進められています。これにより、骨粗鬆症などの代謝性骨疾患の新たな治療法が開発される可能性があります。
さらに、人工知能(AI)を活用した研究も進んでいます。膨大な遺伝子発現データや細胞応答データを解析することで、骨芽細胞の分化を最適に誘導する条件や、特定の患者に最適な骨再生アプローチを予測することが可能になりつつあります。
臨床応用に向けた課題

明治薬品 NMN10000 (60粒/30日分)コエンザイムQ10 NMN 10,000mg 日本製 話題 美容サポート サプリ [シリアルナンバー付]