ヒアルロン酸関節内注射の作用機序から適応疾患、分子量による効果の違いまで医学的エビデンスに基づいて解説。変形性関節症治療における最適な投与タイミングや期間について、最新の研究結果を踏まえた臨床応用とは?
破骨細胞と骨吸収のメカニズムから見る骨代謝疾患
破骨細胞による骨吸収のメカニズムと骨代謝バランスの維持について最新の研究知見をまとめました。骨粗鬆症や歯周病などの骨疾患治療の新たな標的となるシグナル経路とは?

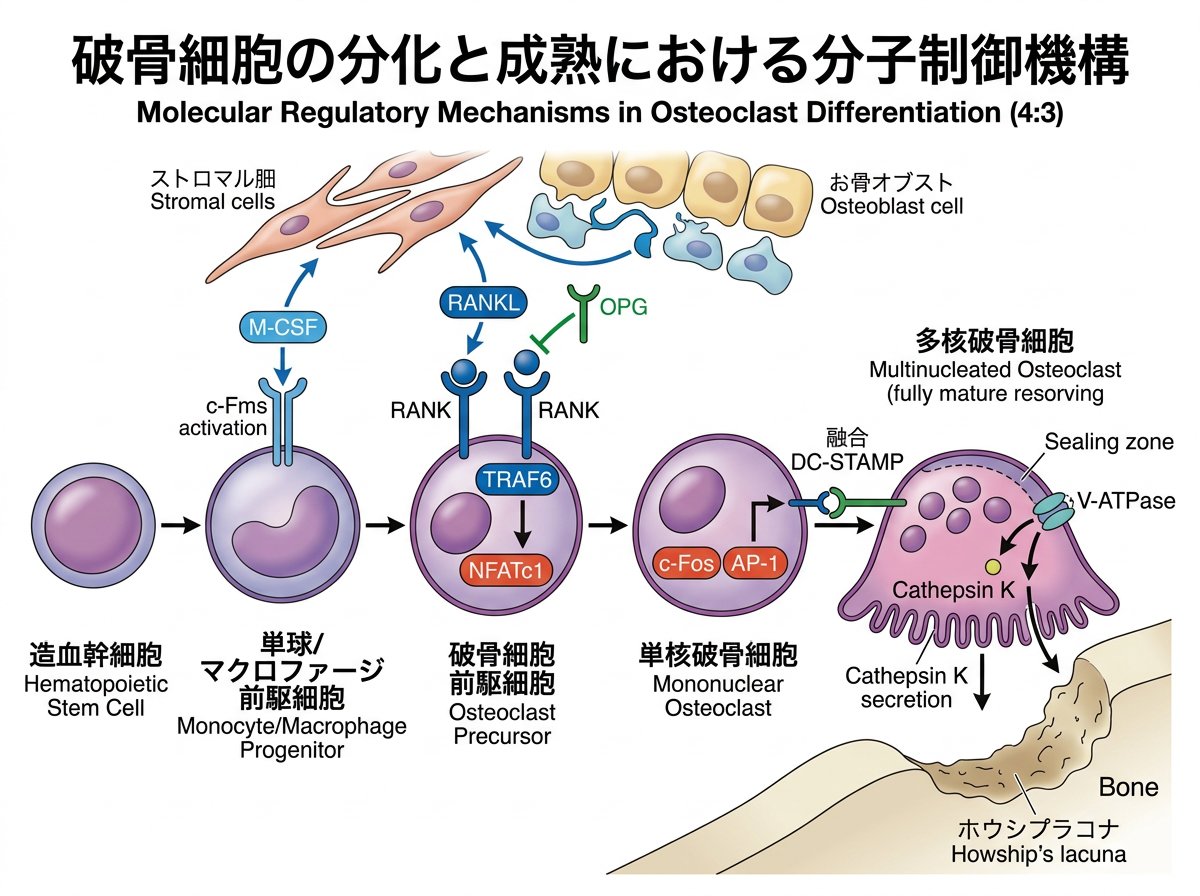
破骨細胞は単球・マクロファージ系の造血幹細胞に由来する多核巨細胞であり、石灰化した骨組織を吸収する唯一の細胞です9。興味深いことに、マクロファージは全身の様々な組織に存在するにもかかわらず、破骨細胞は骨組織にのみ存在します。この組織特異的な分化メカニズムの解明は、骨代謝研究の重要テーマとなっています。
破骨細胞の分化過程は大きく分けて以下のステップで進行します。
この分化過程において、M-CSF (Macrophage Colony-Stimulating Factor) とRANKL (Receptor Activator of NF-κB Ligand) という2つの必須因子が中心的役割を果たします。M-CSFは前駆細胞の増殖と生存を促進し、RANKLは破骨細胞への分化を誘導します。
RANKL刺激を受けると、前駆細胞内でRANK受容体を介して様々なシグナル伝達経路が活性化されます。具体的には、NF-κB、c-Fos、NFATc1 (Nuclear Factor of Activated T-cells, cytoplasmic 1) などの転写因子が誘導され、破骨細胞特有の遺伝子発現プログラムが開始されます。
特に注目すべきなのは、NFATc1が破骨細胞分化のマスター転写因子として機能する点です。NFATc1は酒石酸抵抗性酸性フォスファターゼ(TRAP)やカテプシンKなど、骨吸収に必要な酵素の発現を直接制御します。
また、破骨細胞の形成には前駆細胞同士の細胞融合が必要ですが、この過程には DC-STAMP (Dendritic Cell-Specific Transmembrane Protein) やOC-STAMP (Osteoclast Stimulatory Transmembrane Protein) などの膜タンパク質が関与していることが明らかになっています。
骨組織は一見静的な印象を与えますが、実際には破骨細胞による骨吸収(古い骨の除去)と骨芽細胞による骨形成(新しい骨の合成)が絶えず繰り返される動的な組織です。この「骨のリモデリング」と呼ばれるプロセスは、骨の構造的完全性と品質を維持するために不可欠です。
骨リモデリングの特筆すべき特徴は、骨吸収と骨形成が時空間的に「カップリング」していることです。つまり、破骨細胞が骨を吸収した部位には、続いて骨芽細胞が集まり新しい骨を形成します。この精密な連携が骨量を一定に保つ鍵となります。
カップリングメカニズムとして、以下のシグナル伝達系が同定されています。
このような複雑なシグナルネットワークによって、骨リモデリングの各段階が精密に制御されています。カップリング機構の破綻は骨粗鬆症や骨硬化症などの骨代謝疾患の原因となるため、治療標的としても注目されています。
RANKL-RANKシグナル伝達系は破骨細胞の分化・活性化、そして骨吸収において中心的な役割を果たしています。このシステムの解明は骨代謝研究の重要なブレークスルーとなり、新たな治療法開発への道を開きました。
RANKLは主に骨芽細胞や骨細胞、活性化T細胞などによって産生される腫瘍壊死因子(TNF)スーパーファミリーに属する膜結合型タンパク質です9。一方、RANKはその受容体として破骨前駆細胞上に発現しています。
RANKL-RANK結合のシグナル伝達経路の主な特徴は以下の通りです。
特筆すべきは、この経路を調節するOPG (Osteoprotegerin)の存在です。OPGは骨芽細胞や間質細胞から分泌される可溶性タンパク質で、「おとり受容体」としてRANKLと結合し、RANKとの相互作用を阻害します。つまり、骨吸収活性はRANKLとOPGのバランスによって精密に制御されているのです。
RANKL-RANK系の重要性は、遺伝子改変マウスや遺伝性疾患の研究からも裏付けられています。RANKLまたはRANK遺伝子を欠損したマウスやヒトでは、破骨細胞が形成されず、骨大理石病(大理石骨病)と呼ばれる重度の骨硬化性疾患を発症します。
最近の東京科学大学の研究チームは、NanoBiT技術を用いてRANK-RANKL結合を生きた細胞内で可視化することに成功し、結合部位の特定や定量化を可能にしました。これにより、破骨細胞分化機構のより詳細な理解や、骨粗鬆症治療薬の探索が進展することが期待されています。
RANK-RANKLシグナルの詳細に関する最新研究(日本生化学会誌)
破骨細胞の機能異常、特に過剰な骨吸収活性は様々な骨代謝疾患の発症と進行に密接に関連しています。代表的な疾患と破骨細胞の関わりについて詳細に見ていきましょう。
骨粗鬆症
骨粗鬆症は最も一般的な代謝性骨疾患であり、破骨細胞の活性亢進により骨吸収が骨形成を上回った状態が継続することで発症します。閉経後の女性に多い原因は、エストロゲン減少によりT細胞からのRANKL産生が増加し、破骨細胞が活性化するためです。また、加齢に伴う慢性炎症も骨粗鬆症の増悪因子となり、IL-1、IL-6、TNF-αなどの炎症性サイトカインが破骨細胞形成を促進します。
歯周病
歯周病における歯槽骨破壊のメカニズムは、破骨細胞の活性化が中心的役割を果たしています。歯周病原細菌、特にPorphyromonas gingivalisのリポ多糖(LPS)やペプチドグリカン(PGN)は、宿主の免疫応答を介して破骨細胞形成を促進します。興味深いことに、P. gingivalisが産生するLys特異的ジンジパイン(Kgp)は、破骨細胞抑制因子であるオステオプロテジェリン(OPG)を分解することで、間接的に破骨細胞形成を促進します。また、歯周病では咬合力(メカニカルストレス)も破骨細胞活性化に寄与します。
関節リウマチ
関節リウマチは滑膜の炎症と骨・軟骨破壊を特徴とする自己免疫疾患です。滑膜の炎症部位では活性化T細胞やB細胞、滑膜線維芽細胞からRANKLが過剰に産生され、破骨細胞形成が促進されます。また、TNF-αやIL-6などの炎症性サイトカインもRANKL非依存的に破骨細胞分化を誘導することが報告されています。
骨転移がん
がん細胞が骨に転移すると、「悪性の骨代謝回路」が形成されます。がん細胞はPTHrP(副甲状腺ホルモン関連タンパク質)などを分泌し、骨芽細胞のRANKL発現を増強します。これにより活性化した破骨細胞は骨を吸収し、骨基質内のTGF-βやIGFなどの成長因子を遊離させ、さらにがん細胞の増殖を促進するという悪循環が生じます。
これらの疾患に共通するのは、RANKL-RANK系を中心とした破骨細胞活性化と、骨吸収・骨形成のカップリング機構の破綻です。そのため、これらの分子機構を標的とした治療法の開発が進んでいます。例えば、抗RANKL抗体(デノスマブ)は骨粗鬆症治療薬として広く使用されており、高い骨折予防効果が示されています。
破骨細胞による骨吸収は非常にエネルギー消費の大きいプロセスであり、その活性はエネルギー代謝と密接に関連しています。この破骨細胞特有のエネルギー代謝機構は、新しい治療標的として注目されています。
破骨細胞の骨吸収活性には、プロトンポンプ(液胞型H⁺-ATPase)とクロライドチャネルを利用した塩酸の分泌が必須です。この強酸性環境(pH 4.5程度)によって骨の主成分であるヒドロキシアパタイト(リン酸カルシウム)が溶解します。続いて、カテプシンKなどのプロテアーゼによってコラーゲン線維も分解されます。これらのプロセスを維持するために、破骨細胞は多量のATPを消費します。
破骨細胞のエネルギー代謝の特徴として以下の点が挙げられます。
従来の骨吸収抑制薬(ビスホスホネート、抗RANKL抗体など)とは異なり、破骨細胞のエネルギー代謝を標的とした新規治療アプローチが研究されています。
破骨細胞のエネルギー代謝研究は比較的新しい分野ですが、すでにいくつかの有望な前臨床研究結果が報告されています。例えば、解糖系酵素の1つであるPKM2(ピルビン酸キナーゼM2)の阻害剤は、マウス実験で破骨細胞形成を抑制し、骨量減少を予防することが示されています。
これらのアプローチの利点は、既存の骨吸収抑制薬で見られる顎骨壊死などの副作用リスクが低減される可能性がある点です。また、エネルギー代謝を標的とすることで、より生理的な骨リモデリングの調節が期待できます。
破骨細胞のエネルギー代謝に関する最新レビュー(Journal of Bone and Mineral Research)
破骨細胞研究の進展は、基礎科学の発展のみならず、高齢化社会における骨粗鬆症などの骨疾患治療の新たな選択肢を提供する可能性を秘めています。エネルギー代謝を中心とした新しい視点からの治療戦略開発が、今後の骨代謝研究の重要なテーマとなるでしょう。
[郵送検査キット]骨粗しょう症検査(尿中DPD)【骨粗しょう症リスクチェック】